À la une


Focus recherche
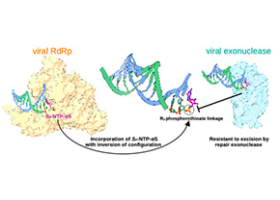
Des analogues de nucléotides qui échappent aux mécanismes de défense des virus
Microbiologie
La biologie pour tous
Dans les poils racinaires, un ballet cellulaire orchestre la fin de la croissance
Biologie végétale


Quatre idées reçues sur les vaccins
Des chercheurs ont mis en évidence un mécanisme de transmission inédit chez les deltavirus. Incapables de se propager seuls, ces virus peuvent se dissimuler à l'intérieur d'un autre virus, appelé virus "helper", qui leur sert à la fois de moyen de transport et de porte d'entrée dans une nouvelle cellule.
À voir aussi
Année thématique
Après l’Année de la Biologie, un dispositif pérenne d’échanges enseignants-scientifiques
Organisée par le CNRS et le Ministère de l’Éducation nationale, de la Jeunesse et des Sports, l’Année de la biologie vise à rapprocher les mondes de l’enseignement et de la recherche pour mettre en lumière les grandes avancées et les enjeux de la…


Un guide pour accompagner l’expression publique des scientifiques du CNRS
Afin d’aider les personnels scientifiques qui le souhaitent dans leurs prises de parole médiatiques, le CNRS leur propose un guide de l’expression publique. Véritable boîte à outils, ce document s'appuie sur les recommandations du Comité d’éthique du CNRS et sur les attentes de ses communautés scientifiques exprimées à l’occasion d’une enquête réalisée en mars 2024.
