Une défense antivirale inattendue dans les tissus vasculaires des plantes
Certains virus sont restreints au système vasculaire des plantes et leur interaction avec leur hôte est encore peu étudiée du fait de la difficulté d'isoler ces tissus. Les scientifiques ont étudié, dans la plante modèle Arabidopsis thaliana, le mécanisme de défense contre le virus de la jaunisse du navet (TuYV) restreint au phloème. Les approches génétiques et de séquençage à haut débit combinées à l'enrichissement en tissus vasculaires ont révélé une nouvelle complexité du jeu entre la défense antivirale par RNAi et sa suppression par la protéine virale P0. Ces travaux sont publiés dans la revue Nucleic Acids Research.
Les plantes emploient l’interférence par l'ARN, ou RNAi pour se défendre contre de nombreux virus. Lors de leur phase de réplication, les virus à ARN passent par un stade ARN double brin (ARNdb) qui est reconnu par la plante comme un signal d’alerte et permet la mise en route de son système de défense qui pourra conduire à l’élimination du virus. Pour la plupart des virus à ARN précédemment décrits, une réponse RNAi antivirale efficace s'appuie principalement sur l’activité de la protéine DICER-LIKE4 (DCL4) pour produire à partir de l’ARNdb la majeure partie des ARN "interférents" antiviraux (siARN) d’une taille de 21-nucléotides. Ces derniers sont associés à une protéine ARGONAUTE (AGO), afin de guider celle-ci vers l’ARN viral, ce qui entraine sa dégradation. Parmi les 10 protéines AGOs de la plante modèle A. thaliana, AGO2, au côté d'AGO1, est considéré comme l’effecteur antiviral majeur dans la lutte contre les virus à ARN.
Certains virus sont toutefois restreints au système vasculaire des plantes supérieures et leur interaction avec leur hôte est encore peu étudiée au niveau moléculaire, principalement du fait de la difficulté d'isoler ces tissus. Comprendre cette interaction devient un enjeu important car ces virus se propagent à un rythme alarmant et entraînent une réduction substantielle du rendement des cultures. L’un de ces virus, le virus de la jaunisse du navet (TuYV) appartenant au genre des Polérovirus code pour une protéine suppresseur du RNAi appelée P0. Cette dernière contre la réponse antivirale en forçant la dégradation des protéines AGOs. Toutefois, la complexité de l’interaction entre le TuYV et la défense antivirale au niveau des tissus vasculaires ainsi que le rôle de P0 dans le contexte d’une infection par le virus doivent encore être démêlés au niveau moléculaire.
Les scientifiques mettent en évidence des caractéristiques nouvelles et inattendues de la défense antivirale au niveau du système vasculaire et de sa manipulation par les polérovirus. Contrairement à la plupart des virus systémiques à ARN, l'ARN double brin du TuYV est principalement clivé par l’enzyme DICER-LIKE2 (DCL2) conduisant à l'accumulation de grandes quantités de siARN viraux d’une taille de 22-nucléotides produits dans le système vasculaire. Ces siARN viraux sont ensuite pris en charge par la protéine ARGONAUTE1 (AGO1), qui s’accumule dans les tissus vasculaires et ceci malgré l’activité de dégradation de P0.
Cette étude démontre également que les siRNA viraux dérivés du TuYV déclenchent la production de petits ARN secondaires à partir des transcrits de l'hôte, une caractéristique qui dépend à la fois de DCL2 et d'AGO1. En effet ni DCL4 générant des siARN de 21 nucléotides, ni AGO2 ne sont capables de stimuler la production de siARN secondaires. En privilégiant la combinaison DCL2/AGO1, les Polérovirus ont la capacité de moduler l’expression des gènes de leur hôte à leur profit. Toutefois, d’autres études seront nécessaires pour élucider le rôle exact de ces siARN viraux secondaires et de leurs cibles au cours de l’infection virale.
Il reste également à déterminer, si les résultats obtenus avec le TuYV peuvent être généralisés à d’autres virus restreints au système vasculaire. Finalement, au-delà des observations faites avec le TuYV, ces résultats suggèrent que le rôle d'AGO1 dans l’activité antivirale pour de nombreuses interactions plante-virus a été précédemment été largement sous-estimé.
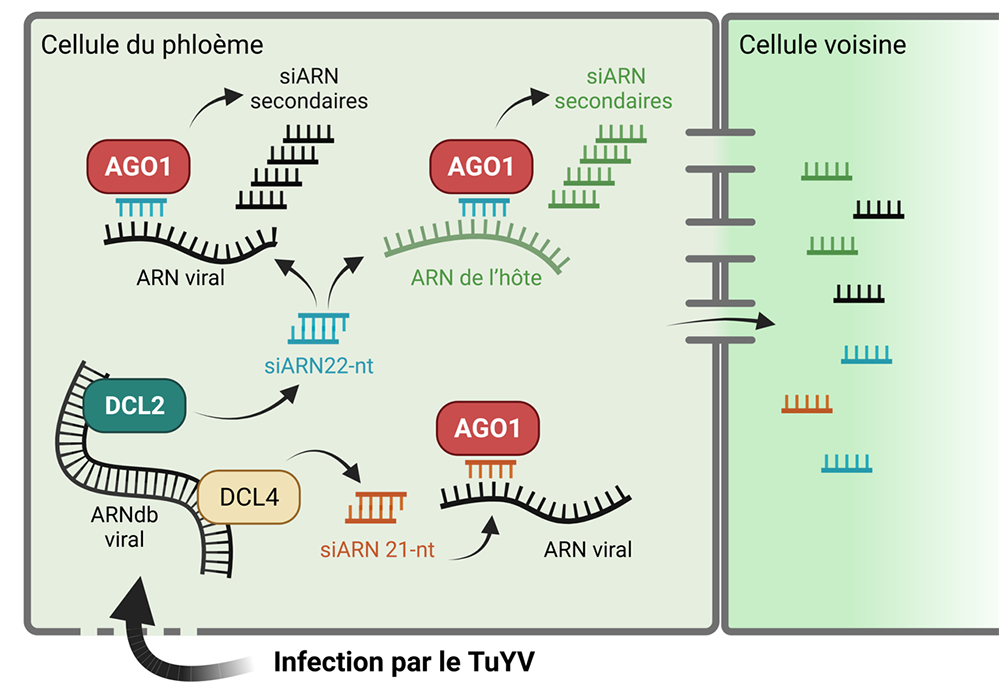
Figure : Schéma illustrant l’infection virale du TuYV dans une cellule du phloème (tissu vasculaire) chez A. thaliana. L’ARN double brin (ARNdb) du virus est clivé à la fois par DCL4 et DCL2 pour produire des siARN antiviraux d’une taille de 21-nucléotides et de 22-nucléotides, respectivement, qui associés à AGO1 permettent le clivage de l'ARN viral. AGO1 associé à des siARN de 22-nucléotides peut également cibler des transcrits de l‘hôte et conduire à la production de siARN secondaires, un phénomène appelé transitivité. Que le ciblage des transcrits de l'hôte par des siARN viraux soit simplement accidentel ou contribue à l'infection et/ou à la réponse de l'hôte nécessitera d'autres investigations.
Pour en savoir plus :
Atypical molecular features of RNA silencing against the phloem-restricted polerovirus TuYV.
Clavel M, Lechner E, Incarbone M, Vincent T, Cognat V, Smirnova E, Lecorbeiller M, Brault V, Ziegler-Graff V, Genschik P.
Nucleic Acids Res. 2021 Oct 6:gkab802. doi: 10.1093/nar/gkab802.
Contact
laboratoire
Institut de biologie moléculaire des plantes – IBMP (CNRS/Université de Strasbourg)
12 rue du général Zimmer
67084, Strasbourg, France