
L’autophagie chez les plantes : découverte des deux enzymes clés pour dégrader et recycler les déchets cellulaires
Comment les plantes libèrent-elles le contenu des vésicules autophagiques une fois celles-ci arrivées dans la vacuole ? En identifiant deux phospholipases complémentaires, LCAT3 et LCAT4, des scientifiques, dans une étude publiée dans Nature Communications, lèvent le voile sur une étape clé du recyclage cellulaire. Cette découverte éclaire le fonctionnement intime de l’autophagie et révèle un rôle inédit de ce processus comme système de transport intracellulaire.
Une étape manquante dans le recyclage cellulaire
L’autophagie est un mécanisme universel chez les eucaryotes, essentiel pour survivre en période de stress (manque de nutriments, sécheresse, etc.). Chez les plantes, comme chez les animaux, ce processus permet de dégrader et recycler des composants cellulaires endommagés ou inutiles, comme des protéines ou des organites, libérant ainsi des ressources et maintenant l’équilibre de la cellule.
Au cours de l’autophagie, les éléments à dégrader sont enfermés dans des vésicules spécialisées appelées autophagosomes. Ces dernières acheminent leur cargaison jusqu’à la vacuole, un vaste compartiment intracellulaire qui assure des fonctions de stockage et de digestion et peut représenter jusqu’à 90 % du volume de certaines cellules végétales.
Si les étapes de formation et de transport des autophagosomes étaient relativement bien comprises, un mystère persistait : comment la membrane de ces vésicules est-elle détruite une fois dans la vacuole afin de libérer leur contenu pour sa dégradation ?
Un mystère résolu : deux phospholipases au cœur du processus
Pour répondre à cette question, les scientifiques ont utilisé la plante modèle Arabidopsis thaliana. En combinant protéomique, génétique, biochimie et microscopie, ils ont identifié deux enzymes de la famille des phospholipases : LCAT3 et LCAT4.
Ces enzymes possèdent la capacité d'hydrolyser les phospholipides constituant les membranes biologiques. Les résultats, publiés dans la revue Nature communications, montrent qu'elles convergent toutes deux vers les corps autophagiques présents dans la vacuole, mais par des voies distinctes.
LCAT4 emprunte directement la voie autophagique. Grâce à son interaction avec la protéine ATG8, acteur central de l’autophagie, elle est incorporée dans les autophagosomes puis transportée à l’intérieur même des vésicules dont elle participera ensuite à la destruction.
LCAT3 suit un itinéraire différent. Elle rejoint la vacuole indépendamment de l’autophagie et semble se positionner sur la face externe des corps autophagiques.
Deux routes, mais une même mission
Une fois dans l’environnement acide de la vacuole, les deux enzymes deviennent actives et agissent de manière complémentaire.
Les données obtenues suggèrent que LCAT3 initie la déstabilisation de la membrane depuis l’extérieur du corps autophagique. Cette première brèche permet alors à LCAT4, confinée à l’intérieur de la vésicule, de poursuivre sa dégradation jusqu’à sa rupture complète. Le contenu peut ensuite être exposé aux enzymes vacuolaires responsables de son recyclage.
Cette coopération explique pourquoi l'absence d'une seule des deux protéines a peu d'effet sur l'autophagie. En revanche, chez les plantes dépourvues simultanément de LCAT3 et de LCAT4, les corps autophagiques s'accumulent dans la vacuole et leur dégradation est fortement ralentie, entraînant une diminution du flux autophagique global.
Les scientifiques ont également confirmé cette complémentarité dans des levures génétiquement modifiées. LCAT3 seule est capable de restaurer partiellement la dégradation des corps autophagiques, tandis que l'ajout de LCAT4 amplifie cette activité.
Une découverte que redessine les rôles de l’autophagie
Ces résultats éclairent une étape méconnue de l’autophagie, essentielle pour :
- la résistance des plantes au stress : sans cette dégradation, les plantes ne peuvent pas recycler efficacement leurs ressources et sont plus sensibles à des carences prolongées.
- L’homéostasie cellulaire : la vacuole, contient des enzymes qui hydrolysent les membranes mais doit protéger sa propre membrane appelée tonoplaste pour éviter de déverser son contenu lytique dans l’ensemble de la cellule.
Ces travaux soulèvent la question de la spécificité spatiotemporelle de LCAT3 et LCAT4 qui, en agissant de manière ciblée sur les corps autophagique, éviteraient une destruction accidentelle de la vacuole elle-même. Dans ce contexte, l’observation que LCAT4 utilise l’autophagie pour rejoindre la vacuole est une découverte centrale qui amène à repenser les fonctions de l’autophagie chez les plantes non seulement comme un processus dégradatif mais aussi un moyen de transport qui pourrait être utilisé par d’autres protéines résidentes de la vacuole.
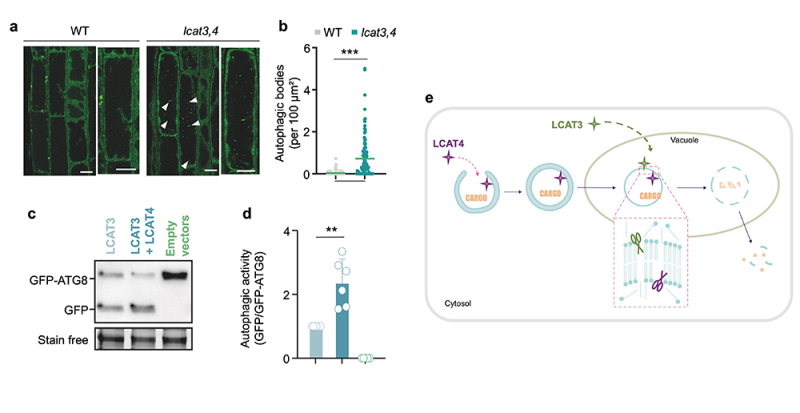
Figure : LCAT3 et LCAT4 désintègrent les corps autophagiques chez la plante modèle Arabidopsis. (a, b) Analyses de microscopie confocale (images en a, quantification en b) de racines de plantules sauvages (WT) ou double mutantes pour LCAT3 et LCAT4 (lcat3,4) montrant l’accumulation de corps autophagiques (flèches) dans la vacuole du mutant en condition de carence en azote et carbone (-NC). Les résultats présentent le nombre de corps autophagiques par 100 μm² de surface de racine et montrent la moyenne et les valeurs individuelles de n=121 (WT) et n=115 (lcat3,4) cellules dans au moins 10 racines indépendantes et 3 expériences indépendantes. p<0.0001, two-tailed unpaired t.test. (c, d) Analyses de l’activité autophagique par western blot suivant la dégradation de GFP-ATG8 et l’accumulation de GFP libre (images en c, quantification en d). Alors que le mutant de la phospholipase Atg15 de levure (S. cerevisiae transformée par les plasmides vides, empty vector) présente une activité autophagique nulle, l’expression d’une version de LCAT3 adressée à la vacuole rétablit partiellement l’autophagie qui est de surcroit augmentée par la co-expression de LCAT3 et LCAT4. Les résultats présentent la moyenne du ratio de GFP/GFP-ATG8 relative à celui de l’échantillon LCAT3, l’écart type ainsi que les valeurs individuelles de n=6 (LCAT3, LCAT3+LCAT4) ou n=5 (empty vector). p=0.006, one sample t.test. . e Schéma récapitulatif du modèle proposé par cette étude.
En savoir plus : Castets, J., Buridan, M., Toboso Moreno, I. et al. A dual phospholipase system instructs membrane hydrolysis during the final stages of plant autophagy. Nat Commun (2026). https://doi.org/10.1038/s41467-026-73116-x
Contact
Laboratoire
Laboratoire de biogenèse membranaire - LBM (CNRS/Université de Bordeaux)
71 avenue Edouard Bourlaux
33140 Villenave d'Ornon